






/ Новости
Развивающиеся части спинного мозга сначала регулируются централизованно, затем переходят на самоуправление
|

Как в ходе развития эмбриона регулируются пропорции его быстро растущих частей — один из актуальных вопросов биологии развития. На примере дорзо-вентральной (спинно-брюшной) организации нервной трубки у мыши и курицы международный коллектив исследователей показал, что способ регуляции может меняться по мере роста органа. Первичное подразделение нервной трубки на участки с разными типами прогениторных клеток, из которых затем образуются разные типы нейронов, определяется градиентами сигнальных белков — морфогенов Shh, BMP и Wnt. Однако в дальнейшем, когда прогениторные клетки начинают дифференцироваться (превращаться в нейроны), влияние морфогенов резко снижается. На первый план выходит автономная регуляция, основанная на характерной для каждого типа прогениторных клеток скорости дифференцировки. Соотношение этих скоростей и определяемая ими дорзо-вентральная организация спинного мозга похожи у мыши и курицы, то есть эволюционно консервативны. Исследование проливает свет на то, каким образом могут сохраняться оптимальные пропорции частей организма при изменениях его размера в ходе эволюции.
Градиенты концентраций сигнальных белков — морфогенов, таких как Wnt, Shh или BMP, играют важную роль в «разметке» развивающегося эмбриона (и отдельных его частей) на дискретные области, клетки которых затем будут вести себя по-разному, формируя разные ткани, органы и отделы тела (см.: Как клетки понимают, что одни должны стать волосами, другие костями, третьи мозгами и т. п.?). Морфогены часто производятся особыми группами клеток (их иногда называют «организаторами», см. Эмбриональная индукция), откуда они распространяются путем диффузии или инициируя собственное производство в соседних клетках, причем концентрация морфогена падает по мере удаления от организатора.
От концентрации морфогенов в межклеточном пространстве зависит, какие наборы регуляторов следующего уровня — транскрипционных факторов — активируются внутри клеток. Транскрипционные факторы, в свою очередь, регулируют экспрессию множества генов, работа которых в конечном счете определяет судьбу клетки.
Градиенты морфогенов, определяющие активацию транскрипционных факторов, — не единственный механизм разметки (patterning) развивающегося эмбриона (см.: Идеи Алана Тьюринга помогли понять механизм развития пальцев у позвоночных, «Элементы», 18.12.2012). Многое в этих механизмах остается неясным. Например, не очень понятно, как регулируются пропорции быстрорастущих частей эмбриона. Ведь рост неизбежно ведет к изменению концентраций морфогенов в развивающихся тканях. Чтобы границы частей при этом не «поплыли», нужно либо динамически менять уровень производства морфогенов, подлаживаясь к меняющимся размерам подконтрольных зачатков, либо использовать другие способы регуляции. Эта проблема относится не только к эмбриональному росту, но и к изменению размеров тела в ходе эволюции. В некоторых случаях, как уже удалось выяснить, действительно происходит подстройка градиентов морфогенов (см.: T. Gregor et al., 2005. Diffusion and scaling during early embryonic pattern formation). Но всегда ли так бывает?
Исследование британских и австрийских биологов, результаты которого опубликованы в последнем выпуске журнала Science, дает отрицательный ответ на этот вопрос.
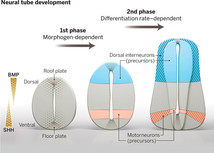
Авторы исследовали регуляцию дорзо-вентральной разметки нервной трубки мышиного эмбриона. Ранние стадии процесса изучены уже достаточно подробно. Крайне упрощенная схема показана на рис. 1, чуть более подробная — на рис. 2.

Разметка контролируется противоположными градиентами двух морфогенов: «вентрального» и «дорзального». На вентральной (нижней, брюшной) стороне нервной трубки, в так называемой донной пластинке (floor plate), а также в нотохорде, лежащем непосредственно под нервной трубкой, синтезируется белок Sonic hedgehog (Shh) — морфоген, участвующий в разметке многих частей организма. На дорзальной (спинной) стороне нервной трубки, в потолочной пластинке (roof plate), синтезируется другой многофункциональный морфоген, BMP (здесь, пожалуй, уместно лишний раз оговориться, что любое описание, тем более популярное, столь сложных биологических процессов — это всегда сильное упрощение, даже примитивизация: в действительности всё, как всегда, намного сложнее).
Таким образом, в нервной трубке создаются два противоположных градиента: сверху вниз растет концентрация Shh, снизу вверх нарастает воздействие BMP. В зависимости от сочетания этих концентраций прогениторные клетки нервной трубки (предшественники клеток нервной ткани) выбирают свою судьбу (cell fate): превращаются из потенциальных предшественников многих разных типов «взрослых» клеток в предшественников только одного типа клеток. Например, моторных или вставочных нейронов. Превращение состоит в том, что в клетках начинает экспрессироваться тот или иной набор транскрипционных факторов. В итоге нервная трубка подразделяется в дорзо-вентральном направлении на 13 доменов (зон), для каждого из которых характерна своя комбинация транскрипционных факторов. В частности, в третьем снизу домене, клетки которого будут в дальнейшем превращаться в мотонейроны спинного мозга, экспрессируется транскрипционный фактор Olig2, а шесть дорзальных доменов, которые дадут начало вставочным нейронам, можно опознать по экспрессии транскрипционного фактора Pax3.
Прогениторные клетки делятся, а потом в какой-то момент некоторые из них начинают превращаться (дифференцироваться) в нейроны того или иного типа. При этом прогениторные клетки остаются в центре, сохраняя контакт с невроцелем (спинномозговым каналом), и продолжают делиться, а нейроны мигрируют на периферию и больше не делятся.
Авторов интересовало, как меняются пропорции прогениторных доменов по мере роста эмбриона и каков механизм регуляции этих пропорций. Первый этап более или менее понятен: первичное подразделение на домены контролируется градиентами морфогенов, а вот что происходит дальше?
Для начала авторы убедились, что рост доменов — резко аллометрический, то есть их относительные размеры по мере роста нервной трубки сильно меняются. Например, домен pMN (область предшественников мотонейронов) сначала имеет довольно большой относительный размер, но потом, на девятый день развития эмбриона, он начинает быстро уменьшаться относительно других доменов (на рис. 2 справа он уже уменьшился). Авторы показали, что размер доменов в точности отражает число прогениторных клеток в них. Самое интересное, что относительные размеры доменов меняются похожим образом и у нормальных мышей, и у карликовых мышей-мутантов, и даже у куриного эмбриона. Значит, динамика пропорций доменов эволюционно консервативна — мало меняется в ходе эволюции. Из этого, в свою очередь, следует, что эта динамика и (или) те факторы, которые ее прямо или косвенно контролируют, очень важны для развивающегося организма.
Что же это за факторы? Авторы рассудили, что число прогениторных клеток в домене может меняться по четырем причинам: 1) пролиферация — деление клеток; 2) апоптоз — программируемая гибель клеток; 3) дифференцировка, то есть превращение прогениторной клетки в нейрон, который покидает домен, сокращая тем самым его размер; 4) смена клеточной идентичности, то есть превращение клетки одного домена в клетку другого домена (для этого должны отключиться одни транскрипционные факторы и включиться другие).
Используя большой арсенал современных методов генной инженерии, окраски и мечения клеток в зависимости от их состояния (наличия тех или иных белков, интенсивности синтеза ДНК и др.), авторы сумели оценить скорости всех четырех процессов на разных стадиях развития нервной трубки.
Скорость деления клеток во всех доменах оказалась практически одинаковой (кроме донной пластинки, где клетки делятся реже). По мере роста она немного снижается, причем это происходит синхронно во всех доменах. Значит, пропорции доменов определяются не темпом пролиферации.
Темп апоптоза оказался пренебрежимо низким по сравнению со скоростью деления клеток. Значит, апоптоз тоже не подходит на роль регулятора пропорций. Он активнее идет у карликовых мышей-мутантов — поэтому спинной мозг у них получается тоньше, но пропорции доменов и в этом случае определяются чем-то другим.
Скорость дифференцировки оказалась разной у разных доменов, и по мере роста она меняется тоже по-разному. Предшественники мотонейронов начинают дифференцироваться раньше других. Максимальная скорость дифференцировки у них тоже достигается раньше (60 hph, hours post headfold stage — через 60 часов после стадии головной складки, у мыши это 10 суток с начала развития). В это же самое время относительный размер домена pMN начинает быстро уменьшаться. В дорзальных доменах темп дифференцировки достигает максимума на 40 часов позже (100 hph).
Дальнейшие тесты и расчеты подтвердили, что различия в скорости роста доменов (а значит, и динамика их пропорций) на поздних стадиях (начиная с 40 hph) объясняются разной скоростью дифференцировки, которая у каждого домена своя. Те домены, в которых дифференцировка идет быстрее, растут медленнее (их относительный размер уменьшается), и наоборот.
Что же касается ранних стадий (<40 hph), то в это время различия в скорости дифференцировки не могут играть решающей роли — просто потому, что дифференцировка еще толком не началась. Чтобы понять, что происходит в этот период, авторам пришлось измерить темпы четвертого из перечисленных выше процессов — смены клеточной идентичности. Для этого использовалась хитроумная методика, позволяющая подсчитать клетки, которые раньше производили определенный транскрипционный фактор, а потом перестали это делать. Были изготовлены генно-модифицированные мыши, у которых, после введения определенного вещества, те клетки, в которых экспрессируется транскрипционный фактор Olig2 (это, напомним, характерная особенность клеток домена pMN — предшественников мотонейронов), начинают производить зеленый флуоресцирующий белок. Если клетка потом перестает производить Olig2 (то есть превращается в клетку другого домена), она всё равно продолжает светиться зеленым. Наличие же в клетке белка Olig2 можно установить при помощи окраски флуоресцирующими антителами (см. Immunostaining). Таким образом, клетки pMN получат две метки одновременно, а бывшие клетки pMN — только одну.
Выяснилось, что темп смены клеточной идентичности — перехода клеток из одного домена в другой путем выключения одних транскрипционных факторов и включения других — высок на ранних стадиях (<40 hph) и резко снижается на поздних.
Как мы помним, включение транскрипционных факторов, маркирующих домены, контролируется градиентами морфогенов. Поэтому данный факт — прозрачный намек на то, что вплоть до 40 hph судьба доменов и их границы определяются морфогенами. В дальнейшем влияние морфогенов снижается, и в дело вступает другой механизм регуляции роста доменов, связанный с темпом дифференцировки.
Чтобы окончательно убедиться в этом, авторы провели ряд экспериментов, в которых они искусственно ослабляли работу сигнального каскада Shh во всех доменах одновременно. Это делалось путем введения ингибитора — циклопамина (вещество получило такое название, потому что, подавляя работу Shh, оно может приводить к циклопии: ведь Shh играет важную роль в том числе и в правильном подразделении лица на две симметричные половинки). Оказалось, что подавление Shh сильно влияет на скорость роста и положение границ доменов до 40 hph, а на более поздних стадиях его эффект резко снижается.
Все эти результаты (а также ряд других, о которых мы умолчали для краткости, включая опыты на эмбрионе цыпленка, показавшие, что у птиц и млекопитающих все эти процессы очень похожи) говорят о том, что регуляция роста доменов осуществляется в два этапа. На первом этапе ведущую роль играют градиенты морфогенов Shh и BMP. Затем чувствительность прогениторных клеток к морфогенам снижается, и рост доменов переходит на «автономный режим» (саморегуляцию): теперь скорость роста и пропорции доменов определяются характерными для каждого домена темпами дифференцировки. От чего зависят эти темпы — следующий важный вопрос, который предстоит разрешить в будущем.
Данное исследование — небольшой, но важный шаг к решению актуальнейшей проблемы биологии: к пониманию того, как генетическая информация, через посредничество сложных процессов самоорганизации в ходе онтогенеза, воплощается в фенотипе. За конкретными деталями конкретных процессов, разгадываемых учеными, постепенно проступают общие принципы. В данном случае показан переход от «общесистемного» управления к локальному, автономному. Первичная разметка идет под контролем системных сигналов — градиентов разлитых в межклеточном пространстве веществ, производимых некими организующими центрами. Но затем размеченные участки развивающегося органа переходят на самоуправление (хотя, конечно, речь не идет о полной независимости от окружающих тканей и внешних сигналов; на поздних стадиях развития нервной трубки рост доменов продолжает отчасти контролироваться такими сигналами — на это есть ряд прямых указаний в обсуждаемой статье).
От себя добавлю, что опыт компьютерного моделирования онтогенеза (см.: Процессы самоорганизации в онтогенезе многоклеточных: опыт имитационного моделирования) помогает понять логику такой двухфазной регуляции. Разметить эмбрион на части при помощи градиентов морфогенов — просто, удобно и эффективно. Но получить желаемый взрослый фенотип (а не что-нибудь бесформенное с торчащими во все стороны ручками-ножками), не отключив своевременно чувствительность клеток к морфогенам и не переведя отделы тела на «саморегуляцию», очень нелегко. По мере роста эмбриона организаторы неизбежно смещаются, расходятся в пространстве, градиенты «плывут», а вслед за ними плывут и границы отделов тела, и весь план строения. Динамически регулировать производство морфогенов и чувствительность клеток к ним, подлаживаясь к меняющимся пропорциям частей тела, — в принципе можно, но это очень непростая программистская задача. Поэтому, моделируя онтогенезы сложных многоклеточных структур, мы обычно используем именно такую двухфазную схему регуляции, которую обнаружили в развитии нервной трубки авторы обсуждаемой статьи.
Источник: Anna Kicheva, Tobias Bollenbach, Ana Ribeiro, Helena Pérez Valle, Robin Lovell-Badge, Vasso Episkopou, James Briscoe. Coordination of progenitor specification and growth in mouse and chick spinal cord // Science. 2014. V. 345 P. 1577.
Источник: http://elementy.ru/news?newsid=432328
Поделиться в Живом Журнале
Стать расой бессмертных – главная эволюционно-историческая задача человечества в III тысячелетии
Имея мышление бессмертных, парадигму бессмертных в качестве мировоззренческой основы, такие люди обязательно реализуют подобные технологии, и мир радикально изменится. Эволюционная ветвь гомо сапиенс в очередной раз сделает крутой вираж и вынесет человечество к невообразимым высотам, туда, где раньше парили только избранные одиночки – бессмертные и боги.
ПодробнееРобот открыл холодильник и принес оттуда пиво
Немецкие разработчики научили гуманоидного робота-помощника TIAGo самостоятельно искать путь к холодильнику, открывать его и приносить пиво. Модульный суперкомпьютер NVIDIA Jetson TX2, служащий зрительным центром робота, позволил ему не только эффективно проложить путь, но и найти пиво запрошенной марки по этикетке.
ПодробнееВ Швеции попытаются создать электронные копии умерших людей
Руководство крупной сети шведских похоронных бюро «Феникс» поставило перед собой амбициозную цель: попытаться создать максимально правдоподобные электронные копии усопших людей.
ПодробнееУчёные из США разработали искусственный аналог глаза
Новое изобретение представили учёные из Школы инженерных и прикладных наук при Гарвардском университете — они создали искусственный глаз, работающий по принципу человеческого.
ПодробнееАмериканцы занялись разработкой реактивных дронов для истребителей
Массачусетский технологический институт по заказу ВВС США занялся разработкой компактных реактивных беспилотных летательных аппаратов, которые можно было бы запускать со стандартного подвеса для ракет под крылом истребителя. Новая разработка получила название Firefly.
ПодробнееToyota представила гуманоидного робота с экзоскелетным управлением
Компания Toyota представила гуманоидного робота T-HR3, управляемого с помощью экзоскелетного контроллера с шлемом виртуальной реальности. Система позволяет оператору управлять движениями робота на месте или передвигать его, а также чувствовать отдачу при взаимодействии с объектами.
ПодробнееЧеловекоподобный робот научился делать сальто
Специалисты Boston Dynamics научили прямоходящего робота Atlas выполнять сальто. Ролик с демонстрацией его новых способностей опубликован на YouTube-канале компании.
ПодробнееРобот-спасатель от Honda: пять «глаз» и 33 степени подвижности
На Конференции по робототехнике в Ванкувере компания Honda представила прототип робота-спасателя E2-DR. У новинки 33 степени подвижности, пять «глаз» и защищенный от пыли и влаги корпус.
ПодробнееToyota представила автомобиль-робот, в салоне которого сразу 2 водительских места
Казалось бы, суть самоуправляемых автомобилей заключается в том, чтобы максимально обеспечить удобство пассажиров и «убрать» из салона водителя, доверив контроль за ситуацией роботу. Вроде бы логичное решение, но вот автоконцерн Toyota думает иначе. Недавно они представили крайне продвинутую версию самоуправляемого авто. Только вот водительских мест в нем аж целых два.
ПодробнееRHP2 - гуманоидный робот, созданный для того, чтобы падать, подниматься и снова падать
Исследователи-робототехники во всем мире тратят безумно большое количество времени и усилий для того, чтобы предотвратить или уменьшить вероятность падения создаваемых ими роботов.
ПодробнееМифы и факты о сверхумном искусственном интеллекте
Станет ли искусственный интеллект лучшим изобретением человечества или же, наоборот, его худшей ошибкой?
Подробнее/ мнения экспертов и членов инициативной группы
- Художник, теоретик искусства, куратор Государственного центра современного искусства (Калининградский филиал)
 Дмитрий Хаметович
Дмитрий Хаметович
БулатовЧлен инициативной группы«В ближайшем будущем гибридные схемы из комбинаций живых и неживых элементов позволят вернуть утраченные или изначально отсутствующие функции. И конечно, заметно усилить их по сравнению с обычными...»
- Доктор философских наук, профессор, главный научный сотрудник Института философии РАН, сопредседатель Научного совета РАН по методологии ИИ
 Давид Израилевич
Давид Израилевич
Дубровский«... этот проект ["Россия 2045"], безусловно, заслуживает всемерной поддержки. Он инициирован молодыми людьми, полными веры в свою высокую миссию. Это яркий акт пассионарности... вызов нашей академической общественности, среднему, сероватому научному сознанию, лишенному порывов вдохновения».
- Автор концепции психонетики, разработчик методологии и методик деконцентрации, корпуса техник активизации сознания и прямой работы с сознанием.
 Олег Георгиевич
Олег Георгиевич
Бахтияров"...«Россия 2045» заставит нас обсуждать очень серьезные вопросы, и это обсуждение приведет к серьезным переформулировкам. Я полагаю, произойдет смещение задач от продления жизни на управляемое развитие человека с выходом за пределы человеческих ограничений..."
- Биофизик, председатель междисциплинарного семинара по трансгуманизму и научному иммортализму РФО РАН, директор по науке компании «КриоРус», член Координационного совета и один из основателей Российского трансгуманистического движения
 Игорь Валентинович
Игорь Валентинович
Артюхов«Как самолет оказался эффективнее птицы, как подводная лодка плавает быстрее, чем кит, так и искусственное тело в какой-то момент догонит и перегонит тело естественное...»
- Доктор физико-математических наук, профессор, заведующий отделом нейроинформатики Центра оптико-нейронных технологий НИИСИ РАН
 Виталий Львович
Виталий Львович
Дунин-Барковский«Для создания искусственного тела нужен хороший мозг, интеллект. А он может быть и искусственным. Воссоздание органов — очень сложная и ресурсоемкая задача. При работе над искусственным интеллектом затраты минимальны, а результаты колоссальны...»
- Руководитель компании «Нейроботикс»
 Владимир Анатольевич
Владимир Анатольевич
КонышевЧлен инициативной группы«Перенос мозга в искусственное тело — более выносливое, более совершенное — единственная возможность человеческой расе остаться на Земле...»
- Доктор биологических наук, профессор, заведующий лабораторией математической нейробиологии Института высшей нервной деятельности и нейрофизиологии РАН
 Александр Алексеевич
Александр Алексеевич
Фролов«Проблема создания искусственной памяти, сохраняющей содержимое естественной памяти индивидуального человека, хотя и является сложной, но представляется разрешимой...»
- Философ, профессор Оксфордского университета, известный своими работами об антропном принципе, основатель (вместе с Д. Пирсом) Всемирной ассоциации трансгуманистов
 Ник
Ник
Бостром«Цифровой путь [бессмертия] – это наша возможность разработать технологию полного копирования мозга, когда мы могли бы создать очень подробную модель конкретного человеческого мозга и воспроизвести ее на компьютере. Тогда мы имели бы потенциал бесконечного существования, создавали бы запасные копии человека и тому подобное...»
- Кандидат физико-математических наук, координатор международного проекта OpenWorm с российской стороны, научный сотрудник лаборатории Моделирования сложных систем ИСИ СО РАН им. А.П. Ершова
 Андрей Юрьевич
Андрей Юрьевич
Пальянов«...Когда мы разгадаем червя – мы поймем жизнь...»
- Руководитель Кластера космических технологий и телекоммуникаций Фонда Сколково, космонавт-испытатель, член Российской академии космонавтики
 Сергей Александрович
Сергей Александрович
Жуков«Я абсолютно убежден в том, что движение «2045» появилось в нужное время в абсолютно нужном месте, потому что верю в великое будущее России, ее подъем после временных трудностей».
- Доцент, кандидат технических наук, профессор кафедры САиУ
 Лев Александрович
Лев Александрович
СтанкевичПервый этап решения проблем бессмертия человека имеет своей главной целью создание нейроуправляемого аватара – гуманоидного робота с человекоподобным скелетом, набором технических мышц и сенсоров.
- Член-корр. РАН, профессор МГУ, заведующий лабораторией «Психология общения и психосемантика» (МГУ)
 Виктор Федорович
Виктор Федорович
Петренко«Возможно, вырабатывая своеобразную систему значений, не привязанную к нашему конкретному миру, с одной стороны, а с другой – разрабатывая изощренные техники медитации и психопрактики, мы выйдем на контакт с возможными мирами на глубинных медитативных уровнях...»
- Доктор философских наук, канд. психологических наук, главный редактор журнала «Историческая психология и социология истории», профессор МГУ.
 Акоп Погосович
Акоп Погосович
Назаретян«Интеллект современного человека – это искусственный интеллект. Естественным осталось только то, что он на белковом носителе, т.е. естествен не интеллект, а мозг...»
- Доктор биологических наук, заведующая лабораторией липидного обмена РНИИ геронтологии
 Елена Владимировна
Елена Владимировна
ТерёшинаЧлен инициативной группы«И не нужно бояться. Проект по созданию носителя для бессмертного разума очень нужен. Конечно, природа рождает умных, талантливых, гениальных людей, но они смертны. Человечество выходит из этого положения — создает книги. И знания так или иначе передаются потомкам. А представьте, гений будет работать вечно!..»
- Доктор медицинских наук, профессор, заведующий лабораторией роста клеток и тканей Института теоретической и экспериментальной биофизики РАН
 Борис Карпович
Борис Карпович
Гаврилюк«Для кожи киборга нужно просто сделать систему питания. А вообще... мы ведь несложно устроены! Есть всего несколько систем: кровеносная разносит кислород и питательные вещества, выделительная выводит отходы. Остальное — рабочие органы. Вначале можно сделать простейший живой организм. А потом более сложные системы...»
/ Мы в социальных сетях:
![]()
/ Сотрудничество:
По вопросам сотрудничества пишите по адресу:
russia@2045.ru
Контакты для СМИ PR@2045.com
 LinkedIn
LinkedIn
 LiveJournal
LiveJournal
 Google
Google
 Я.ру
Я.ру
 Twitter
Twitter
 Facebook
Facebook
 ВКонтакте
ВКонтакте
 Mail.ru
Mail.ru